Membranes and Membrane Transport Flashcards
Biological membranes are about ___% lipid and ___% protein.
Biological membranes are about 50% lipid and 50% protein.

Lipid classification

In most naturally occurring triglycerides, the ____ fatty acid molecule is ____.
In most naturally occurring triglycerides, the central fatty acid molecule is unsaturated.
In solution, fatty acids spontaneously form. . .
a micelle
In solution, glycerolipids spontaneously form. . .
a lipid bilayer
Cholesterol

Cholesterol in membranes

Glycerophospholipid
If there is no head group, it is phosphatidic acid.
If there is a head group it is phosphatidyl x. (example: if x is a choline, it is phosphatidyl choline).

Types of glycerophospholipid

Types of sphingolipid

glucocerebrosides, unlike phosphatidylinositol, are ___.
glucocerebrosides, unlike phosphatidylinositol, are neutral. They do not carry a phosphate group.
Different membranes have different ____.
Different membranes have different membrane lipid compositions. Different leaflets also have different membrane lipid compositions.

Major Routes of Protein Trafficking
Proteins translated in the cytosol may end up in compartments 1, 2, 3, or 4.
Proteins translated in the ER may end up in 5, 6, 7, 8, or 9.
Proteins from outside the cell may end up in compartments 10 or 11.

Signal sequence for cytoplasm
none
Signal sequence for nucleus
Nuclear localization sequence
ex, PAAKKKKLD
The NLS is a short stretch of basic amino acids arginine (R) and lysine (K)
Signal sequence for mitochondria
Mitochondrial localization sequence

Basic localization flowchart

Signal sequence for peripheral membrane proteins
CAAX motif
Must be c-terminal. example: CCIL-Cterm
Signal sequence for integral membrane protein
Transmembrane domain
Must also contain ER signal seq, as it requires ER processing.
SXXS - Hydrophobic span, ~22 amino acids - R and K-rich region, ~6 amino acids
Ecto domain - Transmembrane domain - cytoplasmic domain
Signal sequence for constitutive secretion
Just the ER localization sequence is sufficient to enter the secretory pathway.
Architecture of nucleus

Nuclear transport occurs through. . .
Nuclear pores complexes
Attached is an image showing scanning EM of both sides of a nuclear pore. Note the 8-fold symmetry.

The nuclear localization sequence is a binding site for ____.
The nuclear localization sequence is a binding site for importins.
The nuclear pore proteins (nucleoporins) have long tails that have repeats containing hydrophobic amino acids like phenylalanine. These hydrophobic tails stick together due to the hydrophobic effect, creating a meshwork that blocks the passage of moderate- to large-sized proteins. The importin proteins have hydrophobic amino acids on their surface that can interact with the hydrophobic tails of the nucleoporins, allowing them to “melt” the meshwork and pass through it. By binding cargo proteins with NLS’s, the importins can piggyback proteins through the meshwork.

Signal sequence for peroxisome
SKL = Serine-Lysine-Leucine

Anchoring of peripheral membrane proteins
All of these proteins are considered peripheral membrane proteins, but only the proteins on the inside of the cell would be translated by cytosolic ribosomes. The protein on the outside of the cell would be translated by a ribosome attached to the ER.
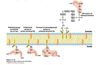
Cartoon of the secretory pathway

Translocon
Channel in the ER membrane that binds proteins by the ER localization sequence as they are being translated and facilitates their translation into the ER.

Signal sequence for non-ER resident proteins that must be processed in the ER.
The signal sequence is cleaved off after successful import in the ER.

Protein disulfide isomerase
Disulfide bonds can only form in the ER and not the cytosol which has a high concentration of reducing agents. Disulfide bridge formation is catalyzed by protein-disulfide isomerase (PDI). It itself becomes reduced in the process and therefore needs to be re-oxidized. The responsible oxidase is Ero1 (ER oxidase 1), which uses molecular oxygen.
If an erroneous disulfide bridge is formed, the protein will get stuck in a high-energy state, most likely partially unfolded, so oxidized PDI may reach and reduce the disulfide bond.

N-glycosylation in the ER

N-glycosylation in the Golgi
When the protein reaches the Golgi, some additional mannose sugars are removed, and other sugars are added. This is referred to as “peripheral glycoslylation”.
NANA=N-acetylneuraminic acid (a sialic acid) Gal=Galactose (a sugar) Man=mannose (a sugar) GlcNAc=N-acetylglucosamine (a sugar) Fucose (sugar)
Note that is therefore possible to determine where a protein is located in the secretory pathway by analyzing the pattern of carboyhydrates attached to the protein.

Retrograde vs Anterograde transport

Constitutive vs Regulated Secretion

Localization of COPI, COPII, and clathrin coats

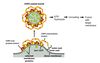
COPII mechanism
The first step is that a protein called Sar1 is recruited to the cytoplasmic face of the ER. Sar1 is a GTP-binding protein. When Sar1 binds to GTP, it exposes a short amphipathic helix that interacts with the membrane. This causes Sar1 to bind to the membrane (yellow protein). Sar1 in turn interacts with other proteins (red), causing them to be localized together into a patch on the ER membrane. These proteins in turn interact with proteins that span the membrane (green). These are the cargo proteins that will be recruited into the vesicle.
In addition, Sar1 binds to proteins that form the outer coat (orange). These proteins help bend the membrane to pinch off the vesicle. The lumen of the vesicle also contains other cargo proteins that bind to the transmembrane proteins (green).
To release upon fusion, Sar1 can hydrolyze the GTP that it binds, and when it does so, it changes shape. This causes the whole coat protein complex to fall apart, leading to a naked vesicle that can fuse with the target membrane.
SNAREs and specific membrane targeting
Fusion is mediated by the SNAREs. Each vesicle and each acceptor compartment contain a SNARE protein (called either v- or t-SNARE for vesicle and target) that pair with each other. Formation of the v-, t-SNARE complex is believed to drive fusion. Notice that the t-SNARE is composed of 3 separate polypeptide chains. The SNAREs all have one domain embedded in the membrane and one domain in the cytosol. The cytosolic domains interact with one another to promote fusion.

Trans vs Cis SNARE states

SNARE Recycling
An enzyme uses chemical energy from ATP to separate the proteins (since the enzyme hydrolyzes ATP in the process it is generically referred to as an ATPase). Once the proteins are separated (lower panel), the V-SNAREs have to be returned to their appropriate compartment so that they can be used again. The T-SNARES can remain where they are, since they are already in the target membrane (by definition)

Receptor mediated endocytosis

Clathrin-coated pits and endocytosis
Clathrin forms coated pits - membrane indentations that are covered on the cytoplasmic side by clathrin. Clathrin can form regular lattice structures that can assemble to form cages. It is thought that the formation of these cages is what drives vesicles to bud from the plasma membrane. After budding the clathrin coat is quickly disassmebled with the help of Hsp70, an ATP driven molecular chaperone that resides in the cytosol.

N-glycosylation for lysosome
For proteins that are trafficked to the lysosome, they are modified differently in the Golgi apparatus. The figure on the left shows the carbohydrate modification of the protein as it arrives from the ER. In the Golgi, and enzyme called GlcNac phosphotransferase uses a molecule of UDP-N acetyl glucosamine to transfer a phosphate to one of the mannose side chains (at position 6 of the mannose, so it is called mannose-6 phosphate). The mannose 6-phosphate is the key tag that helps the protein get sorted to the lysosome.
This means that the enzyme GlcNac phosphotransferase has to know what proteins are bound for the lysosome and which are not. The enzyme only binds to those proteins that are destined to go to the lysosome. Instead of a single short sequence (like an ER signal sequence), the information encoding whether a protein is destined to the lysosome is more complex, and is conveyed in the overall 3D shape of the protein. Those proteins that have the right sequences and shapes to bind to GlcNac phosphotransferase get modified and thus targeted to the lysosome. This method of targeting represents a slightly different concept in trafficking: instead of recognizing only a short sequence motif, this mechanism recognizes a folded shape.
Note that this method therefore also requires that the protein be folded properly before being modified and sorted to the lysosome.

Trafficking of lysosomal enzymes from the Golgi
In the trans-Golgi network, proteins that bear the M6P sorting signal interact with M6P receptors in the membrane and thereby are directed into clathrin/AP1 vesicles (step 1). The coat surrounding released vesicles is rapidly depolymerized (step 2), and the uncoated transport vesicles fuse with late endosomes (step 3). After the phosphorylated enzymes dissociate from the M6P receptors and are dephosphorylated, late endosomes subsequently fuse with a lysosome (step 4). Note that coat proteins and M6P receptors are recycled (2 and 4a), and some receptors are delivered to the cell surface (step 5). Phosphorylated lysosomal enzymes occasionally are sorted from the trans-Golgi to the cell surface and secreted. These secreted enzymes can be retrieved by receptor-mediated endocytosis (steps 6-8), a process that closely parallels trafficking of lysosomal enzymes from the trans-Golgi network to lysosomes.
