Transport mechanisms and renal handling of organic solutes week 1 Flashcards
(13 cards)
What are renal tubules formed by?
T or F: The morphology of cell walls of renal tubules is the same in all parts of the nephron.
For a substance to move across renal tubules, it must either cross ______ or _____ cells.
The renal tubules are formed by epithelial cells. There is different morphology of epithelial cells in different nephron regions as illustrated in Figure 3.1 (attached). This figure emphasizes the point that the renal tubule wall is an epithelial cell layer. It also shows how the characteristics of the epithelial cells vary in different regions of the nephron. For these Physiology lectures, you will not need to know every morphological detail presented here. However, you need to understand that substances must cross a cell layer (either through the cell or btwn cells) in order to move in or out of the renal tubule. This is true at all points along the nephron.

What surface does the basolateral membrane of tubular epithelial cells face? Apical membrane?
The basolateral membrane of a tubular epithelial cell faces the peritubular capillaries and hence the blood stream. The apical membrane of a tubular epithelial cell faces the tubular lumen and for this reason is sometimes called the luminal membrane. The apical membrane may be highly in-folded to increase its surface area. Because of this in-folding, this membrane is sometimes called the “brush border” (usually this term is reserved for the apical membranes of epithelia in the proximal tubule).
What factors does paracellular transport of substances depend on?
Is paracellular transport passive, active, or can it be both?
What substances may be transported via this route?
The paracellular route is the transport pathway where the substance travels between cells. This pathway crosses the so-called tight junctions as well as the intercellular spaces between neighboring cells. Transport via the paracellular route is determined by the permeability (or “tightness” of tight junctions), the concentration gradients across the tight junction and in some cases (i.e. for ions) the transepithelial potential. Paracellular transport is passive and is generally non-selective (at least in comparison to transcellular transport). Examples of substances that may move by the paracellular route are H2O, Cl- and urea.

What factors does transcellular transport of substances depend on?
Is transcellular transport passive, active, or can it be both?
What substances may be transported via this route?
The transcellular route takes the substance through the epithelial cells. This route requires the substance to cross both the apical and basolateral membranes. Substances cross these membranes using a variety of different transport mechanisms (channels, carriers, pumps, etc). Thus, transcellular transport depends on parameters like concentration gradients (across the membranes), membrane potential, membrane permeability and (of course) the presence of particular transporters (channels, carriers, pumps, etc). Transcellular transport can be either passive or active transport. Examples of substances that may move by the transcellular route are H2O, Na+ , Ca2+ and glucose. Transcellular epithelial transport usually requires the cell to be functionally polarized (i.e. transport mechanisms in apical & basolateral membranes are different). For example, the mechanism of Na+ transport at the apical membrane may be different than the mechanism of Na+ transport at the basolateral membrane.

Explain the transcellular transport of Na+ and H2O.
Figure 3.3 (attached) shows an overview diagram of renal transport of salt (NaCl) and water. It presents this transport as a 4-step process. Step 1 is the active transport (directly ATPdependent) of Na+ across the basolateral membrane by the Na-K-ATPase. This moves Na+ up its electrochemical gradient into the interstitium near the peritubular capillaries. The active Na+ pumping lowers the Na+ concentration inside the epithelial cell. Step 2 is Na+ transport across the apical membrane. This may occur via a variety of symporters, antiporters and channels. The driving force for this apical transport is the Na+ gradient that was established by the active Na+ pumping on the basolateral membrane. The net result of steps 1 & 2 (together) is the transport of Na+ from the tubular lumen to the interstitium near the peritubular capillaries. This movement of Na+ (a cation) across the cell layer induces parallel anion transport across the cell layer (i.e. to balance the positive charge moved). Step 3 is the movement of water across the cell layer. The accumulation of Na+ and anions in the interstitial space produces a small local osmotic gradient (between the tubular lumen and interstitium). Water moves down this local osmotic gradient. In essence, water follows the solute. Step 4 is the bulk movement of water and solute from the interstitium into the peritubular capillary.
In Step 2 above, various Na+ dependent transport mechanisms may carry Na+ across the apical membrane. Many of these are symporters that carry Na+ into the cell (down its electrochemical gradient) also carry another solute into the cell. Depending on the particular symporter, this “other” solute could be an amino acid or glucose (among other things). The point here is that Na+ transport is in many cases is linked to the transport of other substances.

What is the site of the reabsorption of most cellular nutrients?
What are 3 generalizations about nutrient reabsorption?
Most common cellular nutrients (e.g. glucose, amino acids, Krebs cycle intermediates, vitamins, etc.) are freely filtered and the proximal tubule is the primary site where reabsorption of these nutrients takes place. The scale of this proximal nutrient reabsorption is huge (as is the filtered load of these nutrients). Some generalizations about nutrient reabsorption can be made and these are listed below.
- Nutrients are usually actively transported (usually by symport with Na+ )
- Most nutrient transport mechanisms have a TM.
- Nutrient transport mechanisms have specificity. Meaning that the transport mechanism will only recognize one (or a few) substances and ignore others. Note: There is not a separate transporter for every substance. Instead, closely related compounds may use the same transport system. For example, a single transporter may handle 3-4 chemically related amino acids. Shared transport pathways mean there may be competition between substances using the pathway. Thus, excess of one amino acid may mean that others will not be transported. This could result in the “others” being excreted.
Explain the reabsorption process of glucose. Indicate whether it is via a transcellular and/or paracellular route.
What is the rate limiting step of glucose reabsorption?
Glucose is freely filtered and all filtered glucose is usually resorbed in the proximal tubule. The steps involved in glucose reabsorption in the proximal tubule are diagramed in Figure 3.4 (attached).
Glucose reabsorption occurs entirely via the transcellular route. The tight junctions in the proximal tubule are nearly impermeable to glucose so no paracellular glucose movement can occur. Renal glucose reabsorption requires a low intracellular Na+ level in the tubular epithelial cell. This is provided by primary active transport of Na+ by the Na-K-ATPase on the basolateral membrane (see Step 1, Figure 3.4). Low intracellular Na+ assures that there is a Na+ gradient across the cell’s apical membrane. The chemical energy in the Na+ gradient is used by the NaGlucose symporter (on the apical membrane) to move glucose up its concentration gradient into the cell from the tubular lumen (see Step 2, Figure 3.4). This is a form of secondary active transport. Once glucose is in the cell, it can passively move out of the cell via the glucose uniporter (GLUT) on the basolateral membrane (see Step 3, Figure 3.4). This is a form of facilitated diffusion. Step 4 is the movement of water and solute from the interstitium into the peritubular capillary. The overall result of this process is transport of glucose from the tubular lumen to the peritubular capillary. In this process, the rate-limiting step is Na-glucose symport (Step 2) and this determines the TM of the glucose reabsorption.
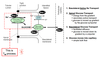
At what plasma glucose level is TM reached? What is the result?
Normally the filtered load of glucose is (0.9 mg/ml 125 ml/min) or 112.5 mg/min. Note 125 ml/min is the normal GFR. Figure 3.5 (attached) illustrates the TM concept for glucose. At a normal plasma glucose level (0.9 mg/ml), the entire filtered load of glucose (112.5 mg/min) will be reabsorbed (see black dotted line) and thus no glucose will be excreted in the urine. If plasma glucose rises to 3 mg/ml (i.e. 300 mg/dL), the renal threshold is reached. This is the point where glucose transport reaches its maximum (TM). If the plasma glucose level goes higher (>3 mg/ml), then the maximal rate of glucose transport will be insufficient to reabsorb the entire filtered load of glucose. In this case, the glucose that is not transported (reabsorbed) will be excreted (i.e. lost in the urine). How the kidneys produce a high volume dilute urine (diuresis) will be discussed later. At this point, however, you may be able to appreciate that if a lot of glucose remains in the tubular fluid, the tubular fluid will be abnormally hyperosmotic and this would tend to draw water into the tubular fluid, resulting in an increased urine volume (or diuresis).

What kind proteins are filtered and must be reabsorbed? Where are they reabsorbed?
Explain the 2 processes of peptide reabsorption. Indicate whether or not there is a TM process.
A common renal mantra is that the glomerular filtrate is protein free. This is not entirely true. The glomerular filtrate is free of most and certainly large proteins but some small proteins (e.g. insulin) and peptides (e.g. angiotensin II) are filtered and these are reabsorbed in the proximal tubule. Some small filtered proteins are transported across the apical membrane by endocytosis. This endocytosis is triggered by the binding of the protein to receptor sites on the apical membrane. Since these sites can saturate, there is a TM for this process. Endocytosis generates a membrane vesicle (containing the filtered protein) in the cytoplasm of the cell. These vesicles merge with lysosomes that contain enzymes that will degrade the filtered protein into small fragments and eventually into amino acids. These then exit the cell across the basolateral membrane and enter the peritubular capillaries. Other small filtered peptides (really small ones) are handled differently but with the same end result (catabolism of the peptide and conservation of its amino acids). The difference is that they are catabolized in the proximal tubular lumen by peptidases that are tethered to the apical membrane. The resulting amino acids are then transported across the apical membrane by the Na-dependent amino acid symporters that normally reabsorb filtered amino acids.
Remember, there is normally almost no protein in urine. Protein in the urine is sign of serious renal dysfunction or disease.
Explain the steps involved in the secretion of organic anions and cations.
Is it via the transcellular or paracellular route? Is there a TM process?
Where in the nephron does secretion of organic anions and cations occur?
Secretion of organic anions: The proximal tubule secretes a wide variety of organic anions. These may be endogenous or foreign substances. Figure 3.6 (attached) shows a table listing some examples. Many of these substances are filtered and thus their secretion adds to what will eventually be excreted in the urine. However, other may be bound to large plasma proteins and thus little will be filtered. In this case, their secretion is the primary pathway to their eventual excretion. Since the tight junctions are typically not permeable to organic anions, the transport pathway for organic anions is the transcellular route. Generally, there are active transporters for these anions on the basolateral membrane that pump these substances into the cell. Then, their transport across the apical membrane is accomplished by facilitated diffusion using uniporters or occasionally by Na-dependent antiporters. The relatively nonselective nature of these transporters accounts for their ability to handle so many different substances. Since these transport mechanisms can saturate, there is a TM for organic anion transport. If the plasma level of an organic anion (say penicillin) is high (above TM), then not all of it will be cleared from the plasma (this is the goal of a penicillin dose regimen).
Secretion of Organic Cations: The proximal tubule also contains several closely related transporters for organic cations (analogous to those for the anions described above). Like the anions, different cations compete for the same transporters and there is a TM for cation transport. Proximal secretion of organic cations (and anions for that matter) is critically important for those cations (and anions) that are bound to large unfiltered proteins. It is important because if the substance is not filtered than secretion is the only other way into the renal tubule. Like the anions, some organic cations are not bound to proteins and thus are filtered (and subsequently secreted as well). The prime example here is the cation creatinine.

Explain the process of urea recycling. Indicate the parts of the kidney where urea is filtered, secreted, and reabsorbed.
What is urea recycling important for maintaining?
Urea is a special substance. It is both a waste product and key to controlling the body’s water balance. Urea is produced continuously by the liver as an end product of protein metabolism. Its rate of production may vary with diet but it never stops. Toxic plasma levels of urea result in a condition called uremia. Over days and weeks, renal urea excretion normally matches hepatic production. This may not be the case over hours and days because urea excretion is regulated for other purposes (other than eliminating what is produced by the liver). Urea is a small, water soluble, freely filtered solute. About 50% of filtered urea is reabsorbed in the proximal tubule. Then, a similar quantity of urea is secreted back into the tubular fluid in the loop of Henle. Then, urea is reabasorbed again from the medullary collecting duct. The net result is that about half of the filtered urea ends up in the urine. The numbers here can (and do) vary with body hydration status. Also note that the last two steps of renal urea handling (underlined above) both occur in the renal medulla. Urea is essentially “recycled” through the renal medulla. Let’s follow a urea molecule through the renal medulla to illustrate the point. A urea molecule is secreted into the loop of Henle. This same urea travels (through the nephron) to the medullary collecting duct where it is reabsorbed. The very same urea may then be secreted (again) into the loop of Henle and do the same cycle again. This urea recycling is important in maintaining body water balance and this will be discussed more later. However, our focus now is only on how urea (an organic solute) is actually handled by the kidney.
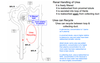
Explain the route of urea reabsorption in the proximal tubule and collecting duct.
Explain the route of urea secretion in the Loop of Henle.
Explain why each route is used in the respective parts of the kidney.
Proximal Tubule Urea Reabsorption
- Na is reabsorbed with H20 following. As H20 leaves tubule, urea is concentrated. This creates a urea gradient across tubule.
- Urea passively diffuses down this gradient along the paracellular route.
- Tight junctions are not so tight.
Urea Transport in Loop & Collecting Duct
- Tight junctions are tight (paracellular not available)
- Urea is transported along transcellular route via facilitated diffusion (urea uniporter)
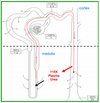
What drives urea secretion in the loop of Henle? What drives urea absorption in the collecting ducts?
Explain how urea levels correlate with hydration status.
During periods of water excess, medullary urea concentration levels off at 300-400 mM (see attached). Nevertheless, the medullary urea level is substantial (compared to normal plasma). In the loop of Henle, the urea concentration is higher than normal plasma but lower than in the renal medulla (i.e. medullary interstitium). Thus, there is a driving force for urea to move down its concentration gradient into the tubular lumen. Uniporters in the loop of Henle facilitate urea secretion into the tubular lumen. As the tubular fluid moves into and through the collecting duct more water is reabsorbed so the urea concentration in the tubular fluid becomes even higher. So high that when it reaches the medullary collecting duct there is a driving force for urea to move down its concentration gradient into the medullary interstitium. Uniporters in the collecting duct facilitate this process and consequently urea is reabsorbed back into medullary interstitium. Since urea levels in the interstitium can vary with body hydration status, the driving forces for medullary secretion (in loop of Henle) and reabsorption (in collecting duct) will also vary with hydration status. Renal handling of urea is complex but the net result (over the long run) is that urea excretion will end up matching hepatic urea production. The body stays will stay in urea balance.

